Emily Wheater // A Design for Life, an exhibit on at the Surgeons’ Hall Museums in Edinburgh, is tiny. Yet, in one room, the contribution of 18th – 19thC comparative anatomy to medical education and training of the day and to the development of early evolutionary theory is told cogently and precisely. The tale encompasses Aristotelean origins with observations of wound healing in animals, a rebirth on the dissection table under Vesalius’ eye, and the establishment of comparative anatomy as a modern scientific discipline in the 18th/19th centuries and the development of early evolutionary theory. The most prominent objects on display are vertebrate skeletons, some replicas and some real, some partial and some whole. They illustrate both shared body plans and how individual species are characterised by their adaptations of this shared body plan to their individual niches and environments. The forelimbs of a bat, a mole, and a human have the same components, even if the final products are vastly diverging. A contrast is drawn between the streamlined, fragile-looking skull of a gannet and the heavy, vegetation crushing mass of a capybara skull. It is through juxtapositions like these that the exhibition offers a deft demonstration of the principle that structure relates to function, a keystone of biology.
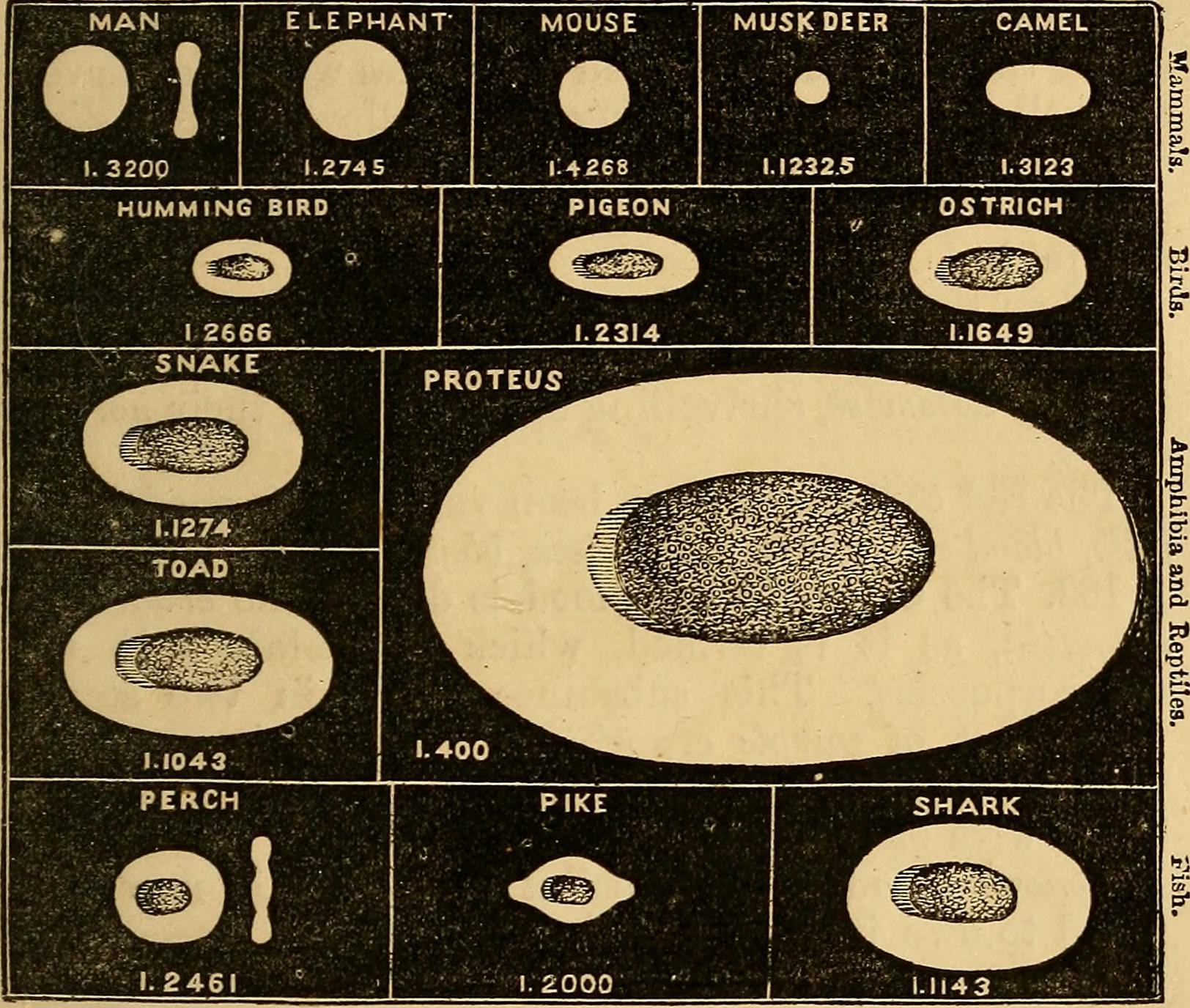
In one cabinet, placed on the very bottom level, is a seemingly humble and very beige collection of objects. These were wax models of erythrocytes – red blood cells – from several different species, including frog, deer, and human (two models for human). They were circular or elliptical, with that distinctive doughnut dip in the middle, and more varied in size than I would have expected. The cells were represented at 5000x magnification and were notable, in an exhibition that demonstrates so well the importance of macro level anatomy to the scientists and medics of the 19thC, for the objects of interest being microscopic. Microscopic comparative anatomy is still significant today, especially in the domain of biomedical research. As a Masters research student I worked in a research group using zebrafish as a model organism for studying sleep. When I told friends and family members about my project, the first question that was inevitably asked was “do zebrafish sleep?” The answer would involve a discussion of shared sleep-wake behaviours, but also of neural circuit and neuropeptide conservation of sleep regulating pathways between zebrafish and human [1] [2]. In other words, evidence of analogous neural systems, gained from microscopic comparative anatomy, supports the use of zebrafish as an informative and valid model for studying the biological process of sleep, even though they look so very different from humans. The evidence looks different now; we have progressed from wax models of red blood cells, or diagrammatic representations of the concordance of knee joints, to images captured by cameras mounted on microscopes, often fluorescence microscopy or in situ hybridisation techniques [3]. Despite this difference in presentation, the logic underlying the work is shared.
In modern times the animal of choice for biomedical research is the mouse, followed by the zebrafish (in 2017 the UK Home Office published statistics that showed that for ‘Translational/Applied Research’ 59% of animal procedures had been carried out on mice compared to 19.5% in the zebrafish) [4]. Mice, being mammals, are appealing due to their phylogenetic closeness and physiological similarities to humans, criteria of ‘relevance’ that owe their existence to evolutionary biology. But evolutionary ‘closeness’ is just one facet of judging how an animal model may or may not be relevant to a human disease or condition. Mice are small, nocturnal, and short-lived. Their diets and behaviours are different from those of humans. This becomes particularly troublesome when using them as models for human conditions where the etiology is poorly understood, such as psychiatric diseases. Mouse models of depression, for instance, may involve the removal of the olfactory bulbs from the brain, or a series of mild stressors applied over a long time period. The latter (usually termed the ‘chronic mild stress model’) is thought to bear etiological validity to the human condition in a way that removal of the olfactory bulb is not [5]. But to which species, in this model scenario, do we owe ecological validity? Should we try to induce human depression in mice, or should we study a depression that is perhaps more endogenous to the mouse? It is far from clear which approach is more likely to provide us with the novel anti-depressant strategies that are ultimately the purpose of this research.
Research efforts in animal models are predicated on assumptions of similarity and concordance, that studying what occurs in one species is informative about what occurs in another. But, as A Design for Life demonstrated, there are two prongs to comparison. One is the principle of interconnectedness and similarity: shared body plans, analogous processes and systems. The other is that of difference and distinctions, manifested in species-specific adaptations and formalised into taxonomical boundaries. It is in relation to these principles, established by Enlightenment period thinkers, that we still frame the justifications and the limitations of seeking answers about human biology from the study of other species – a bedrock of modern-day biomedical science.
References
[1] Kaslin, J. & Panula, P. Comparative anatomy of the histaminergic and other aminergic systems in zebrafish (Danio rerio). J. Comp. Neurol. 440, 342–377 (2001).
[2] Zhdanova, I. V. Sleep and its regulation in zebrafish. Rev. Neurosci. 22, 27–36 (2011).
[3] Liu, J. et al. Evolutionarily conserved regulation of hypocretin neuron specification by Lhx9. Development 142, 1113–1124 (2015).
[5] Harkin, A., Kelly, J. P. & Leonard, B. E. A review of the relevance and validity of olfactory bulbectomy as a model of depression. Clin. Neurosci. Res. 3, 253–262 (2003).
Image credit: From “Elements of animal physiology, chiefly human” (1873) – https://www.flickr.com/photos/internetarchivebookimages/14596314327/in/photostream/
A Design for Life is currently on at the Surgeons’ Hall Museums in Edinburgh, until Easter 2020.



{kind=link}